

![]()
Abstract
The
extended-spectrum-β-lactamases (ESβLs) are enzimes that can
hidrolize the amide ciclic bond of the oxyminos-β-lactamics (cefotaxime,
ceftazidime and aztreonam). These are encoded in megaplasmids of more
than 80 kDa, that generally posees others genes of antimicrobial resistance.
Recently, there have been described another genetic structures, called
integrons, through which different genes can be stored and disseminated
between strains. They posees the genetic information requiered to express
a protein (integrase), implied in the capture and storing of antimicrobial
resistance genes. In this investigation we detected class 1 integrons
in Enterobacterias that produce ESβLs. We studied a sample of 51
Enterobacterias isolated from seven hospitals of Caracas-Venezuela.
The fenotipic and molecular detection of ESβLs was realized, in
the following, plasmids were transfered by conjugation in a solid media
and isolated from donor and transconjugant strains. Finally class 1
integrons were detected by PCR, employing primers for the class 1 integrase.
The 68.6% of the strains present ESβLs type SHV, 15.7% type CTX-M-group
2, and 7.8% carry both types of ESβLs. From 36 conjugated strains,
81% transfered plasmids carriers of genes that coded ES?Ls. The analysis
of the plasmidic isolates, showed in all the transconjugant strains,
a band of 25.000 pb and in a 80% a plasmidic band larger than 50.000
pb was observed. The 27.5% (14) of donor strains and their respective
transconjugants are carriers of class 1 integrons asociated to conjugative
plasmids. The 32.4% of the strains with integrons produce ESβLs
type SHV, instead the 44.4% produce ESβLs type CTX-M.
Introducción
La rápida aparición y diseminación de microorganismos resistentes a los agentes antimicrobianos, constituye un problema de salud pública a nivel mundial. La presión selectiva ejercida mediante el uso excesivo de antibióticos, junto a los diversos mecanismos de transferencia genética que portan las bacterias, contribuyen cuantiosamente con esta situación.
En la actualidad, los β-lactámicos siguen constituyendo una de las mejores opciones terapéuticas antimicrobianas. Sin embargo, la efectividad de los mismos ha sido contrarrestada por la aparición de las β-lactamasas. Los genes que codifican estas enzimas pueden residir en el cromosoma bacteriano o en plásmidos8. Estas últimas de mayor relevancia ya que pueden ser transferidas a diversos géneros bacterianos y diseminarse en el ambiente hospitalario y en la comunidad.
Las
β-lactamasas de espectro expandido (BLEE) son enzimas que presentan
un espectro incrementado de actividad hidrolítica, contra las
cefalosporinas de tercera y cuarta generación y sobre el aztreonam.
Son derivadas a partir de mutaciones puntuales de las β-lactamasas
de espectro ampliado (BLEA) TEM-1, TEM-2 y SHV-117, aunque
existen otras familias de BLEE, como las tipos CTX-M y PER que tienen
orígenes diferentes y una escasa relación estructural
con las TEM y SHV.
La movilización y estabilización de genes de resistencia
a antibióticos ha sido conducida por los mecanismos de transferencia
horizontal de la información genética, principalmente
a través de la conjugación1, 2, 6. La transferencia
de ADN mediante conjugación, esta mediada por plásmidos
conjugativos, que contienen toda la información genética
necesaria para promover su propia transferencia1.
Plásmidos transmisibles y/o transposones permiten el intercambio de genes que codifican para determinantes de resistencia a múltiples antibióticos, entre especies poco relacionadas, representando el medio más común para adquirir determinantes de resistencia a antibióticos.
En años recientes se han reportado la existencia de otra clase de unidades genéticas denominadas integrones. Estos, son elementos pertenecientes a la familia de transposones Tn21 y Tn3 1, capaces de albergar genes de resistencia a antibióticos. Los integrones están constituidos por dos regiones de ADN muy conservadas, situadas en sus extremos. En la región conservada 5´ (5´CS) existen 3 elementos necesarios para la captura y expresión de genes exógenos (cassettes): uno que codifica una integrasa (int I), el otro es el lugar de recombinación sitio-específico (attI), y un promotor (Pant) para la expresión de los genes cassettes integrados. En la región conservada 3´ (3´CS) se hallan insertados el gen qacEA1 y el gen sulI 11. Entre estas dos zonas se pueden insertar uno o varios tipos de genes (casettes génicos) de resistencia, en una zona denominada región variable 15 (Figura 1).
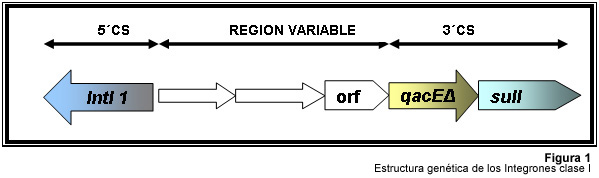
De acuerdo con la secuencia de la integrasa se han clasificado los diferentes tipos de integrones. Se conocen nueve clases, los clase 1, 2 y 3 contienen genes de resistencia antimicrobiana. Algunos genes de β-lactamasas pueden estar codificados en dichos cassettes génicos 9, 19.
Los integrones no son móviles por si mismos, pero con frecuencia se hallan asociados a transposones que a su vez se encuentran en plásmidos conjugativos, por lo que su movilidad horizontal esta asegurada 21.
Los integrones aparecen como estructuras genéticas que albergan genes de resistencia denominados cassettes génicos, permitiendo de esta manera el reconocimiento, la captación, intercambio y almacenamiento de nuevos mecanismos de resistencia, como es el caso de las β-lactamasas. En Venezuela son escasas las investigaciones que asocian la presencia de integrones clase 1 en bacterias multirresistentes aisladas de ambientes hospitalarios. Además, no existen en nuestro país reportes de la presencia de BLEE asociadas a este tipo de integrones.
Es por ello la importancia de la identificación de integrones, especialmente en cepas de origen nosocomial, en las cuales la multirresistencia puede estar asociada a estas estructuras.
En esta investigación se demostró la presencia de integrones clase I, asociados a plásmidos conjugativos in-vitro, en cepas de Enterobacterias y sus respectivas transconjugantes productoras de BLEE de las familias SHV y CTX-M.
![]()
![]()