
![]()
Óxido nítrico
El interés por el NO como transmisor neuronal surgió con el hallazgo
experimental de su liberación por las células granulosas del cerebelo
cuando son expuestas a la acción de sustancias agonistas del glutamato4.
El NO, identificado inicialmente con el nombre de factor de relajación
derivado del endotelio5, es un gas con estructura de radical libre,
extremadamente lábil y ligeramente soluble en diversos solventes, que
difunde con relativa facilidad a través de las membranas biológicas.
La molécula de NO es sintetizada a partir de su precursor biológico, el aminoácido L-arginina, mediante la acción de la enzima sintetasa de óxido nítrico (NOS, por sus siglas en inglés, EC 1.14.13.39), la cual posee al hemo como grupo prostético responsable de la catálisis oxidativa y cuenta además con sitios de reconocimiento funcional para diversos donadores de electrones. Existen por lo menos tres isoformas de la NOS, de las cuales dos son constitutivas (NOSe, descubierta en el endotelio de los vasos sanguíneos, y NOSn, descubierta en las células nerviosas), y una inducible (NOSi) mediante un estímulo inmunológico. Las células que contienen las isoformas constitutivas producen NO en cantidades relativamente bajas y por cortos períodos de tiempo. En cambio, cuando la isoforma NOSi es inducida, son producidas grandes concentraciones de NO. La acción catalítica de las enzimas constitutivas es dependiente de calcio y calmodulina, mientras que la de la enzima inducible no muestra esta dependencia aún cuando posee sitios de unión tanto para calcio como para calmodulina 6. En el sistema nervioso, la mayoría de las neuronas expresa primariamente la isoforma NOSn, si bien algunas neuronas también expresan la isoforma NOSe, aunque en diferentes proporciones, y, bajo ciertas condiciones, pueden ser inducidas para que expresen la forma inducible de la enzima.
En el cerebro, la localización de las neuronas que contienen NOSn es variada y extensa pero con una mayor proporción en el bulbo olfatorio y en las células granulosas del cerebelo.7 Por otra parte, con técnicas de inmunohistoquímica y de hibridización in situ, se ha demostrado que en el cerebro la enzima NOSn tiene una distribución que no se correlaciona con ningún otro neurotransmisor 8,9. Así, en el cerebelo, la presencia de NOSn ocurre en las células granulosas glutaminérgicas y en las células en cesta GABAérgicas; mientras que, en la corteza cerebral, las neuronas que presentan NOSn son células que contienen somatostatina, neuropéptido Y, y ácido y-aminobutírico (GABA), y en el cuerpo estriado, la colocalización se presenta en neuronas que contienen somatostatina y neuropéptido Y. En el hipocampo, la NOSn se localiza solamente en las interneuronas GABAérgicas; mientras que, en las células piramidales de la región CA1-CA3, no observan cantidades detectables de NOSn sino de NOSe, como ocurre también en las células granulosas del giro dentado.10
En el cerebro de los humanos el NO es producido mediante la oxidación enzimática del grupo guanidina del aminoácido L-arginina para formar citrulina y NO con la producción intermedia de hidroxi-L-arginina. La reacción es dependiente del complejo calcio-calmodulina y tiene lugar en presencia de oxígeno y con la participación de tetrahidrobiopterina como cofactor y de la forma reducida del fosfato dinucleótido de nicotinamida adenina (NADPH) como donador de electrones. La unión del complejo calcio-calmodulina a la NOSn es esencial para la producción de NO. La NOSn es activada por el complejo calcio-calmodulina después de que se produce un incremento en los niveles intracelulares de calcio provocado por la acción de aminoácidos excitadores como el glutamato.11 Cuando el glutamato liberado por la neurona presináptica activa al receptor N-metil-D-aspartato (NMDA) de la neurona postsináptica el calcio entra al citosol, se une a la calmodulina y activa a la NOSn, la cual se encuentra físicamente unida al receptor NMDA por la proteína de densidad postsináptica-95 (PSD-95).12 Esto proporciona una ruta directa para el calcio a través de los receptores NMDA que le permite alcanzar la NOSn casi inmediatamente (Fig. 1).
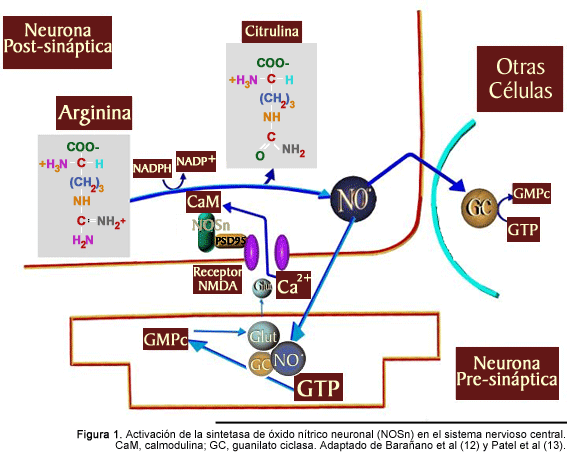
El NO no se almacena en la neurona postsináptica y su liberación se lleva a cabo por difusión simple hacia la neurona presináptica o a células vecinas (neuronas o astrocitos) para interactuar con la enzima guanilato ciclasa soluble para producir GMPc 13. El NO se une al hierro en el centro activo de la guanilato ciclasa para iniciar un cambio conformacional en la enzima, con lo que se incrementa la actividad del dominio ciclasa, y, por consiguiente la producción intracelular de GMPc. Los mecanismos exactos por los cuales el GMPc ejerce sus efectos biológicos son poco conocidos. No obstante, se sabe que el GMPc actúa como segundo mensajero y como tal puede estimular la liberación de glutamato, afectar la función de canales iónicos, facilitar la fosforilación de varias proteínas, o activar quinasas proteicas que intervienen en diversos eventos celulares. El NO producido en las neuronas también es capaz de difundir a los vasos sanguíneos cercanos y producir vasodilatación. Sin embargo, no todos los efectos del NO son mediados por el GMPc, ya que el NO puede interactuar con el oxígeno molecular o el radical superóxido para producir especies reactivas de nitrógeno que pueden modificar diversas macromoléculas incluyendo proteínas, lípidos y ácidos nucleicos14
La función precisa del NO en las neuronas permanece sin ser aclarada plenamente. Hay efectos directos e indirectos. Con bajas concentraciones (<1µM), predominan los efectos directos, es decir, aquellos que son mediados por la molécula de NO. Con altas concentraciones (>1µM), prevalecen los efectos indirectos del NO mediados por las especies reactivas de nitrógeno. En otras palabras, la concentración, y por lo tanto la fuente de NO, son los principales factores determinantes de los efectos biológicos del NO.15
Sin embargo, la amplia distribución de las neuronas que contienen NOSn en el sistema nervioso central hace pensar que el NO podría estar involucrado en numerosas funciones. Una de las funciones atribuidas al NO se relaciona con la morfogénesis del sistema nervioso.16 Durante el desarrollo, el NO puede influenciar el establecimiento de la organización columnar de la corteza cerebral en vista de que, entre los días 15 y 19 de la gestación, la expresión de la NOSn se presenta selectivamente en la placa cortical. Asimismo, en el bulbo olfatorio se observa la presencia de la NOSn desde el día 15 hasta etapas tempranas de la vida postnatal. Esto ha llevado a proponer que el NO puede funcionar como factor trófico para el desarrollo de las neuronas, y determinar además cuales van a entrar en apoptosis. El NO también ha sido involucrado en los cambios que subyacen a la memoria y al aprendizaje 17,18,19 La atención se ha enfocado en algunas formas de plasticidad sináptica como la potenciación de larga duración en las células piramidales CA1 del hipocampo (cuyo correlato in vivo parece ser la memoria), donde la estimulación presináptica submáxima induce la liberación postsináptica de NO, el cual actúa en el terminal presináptico para incrementar la liberación del neurotransmisor y, por consiguiente, la respuesta postsináptica. Adicionalmente, el NO ha sido involucrado en el proceso de transducción de varias modalidades sensoriales. En la vía visual, por ejemplo, el NO interviene en diversos procesos que van desde la transmisión sináptica de los bastones con las células bipolares hasta el control de la excitabilidad neuronal en la corteza visual.20 En los órganos neuroendocrinos, la localización de las neuronas NOSn cercana a las neuronas productoras de hormonas, como ocurre con las neuronas productoras de la hormona liberadora de gonadotropinas (GnRH) en el hipotálamo, sugiere que el NO podría mediar una variedad de importantes funciones neuroendocrinas.21
En el sistema nervioso periférico, el NO actúa como neurotransmisor en la fibras nerviosas no adrenérgicas no colinérgica (NANC) para mediar algunas funciones gastrointestinales y genitourinarias. Así, por ejemplo, el NO es el mediador en la relajación de los músculos del fundus gástrico que se produce cuando son estimulados sus nervios intrínsecos 22; en el intestino, el NO difunde de las neuronas del plexo mientérico hacia las células de músculo liso adyacentes para provocar su relajación 23; y, en el aparato genito-urinario, el NO es el neurotransmisor de los nervios que inervan los cuerpos cavernosos del pene. La estimulación de estos nervios permite la erección del pene.24
Por otra parte, la sobreproducción de NO en el cerebro puede tener un papel en la patogénesis de ciertos trastornos neurodegenerativos y en algunas formas de neurotoxicidad. La destrucción neuronal que se presenta en los casos de excitotoxicidad puede ser desencadenada por un exceso de glutamato en el espacio extracelular.25 La estimulación prolongada de los receptores NMDA por parte del glutamato provoca el ingreso masivo de calcio al interior de la célula con la consiguiente sobreproducción de NO, el cual, una vez liberado, estimula la liberación de glutamato en las células vecinas al tiempo que sensibiliza a los receptores NMDA a la acción excitatoria del glutamato. Por otra parte, el incremento en la entrada de calcio hasta alcanzar niveles citotóxicos desencadena una sucesión de reacciones químicas mediadas por enzimas proteasas, endonucleasas y fosfolipasas que provocan, entre otras cosas, la desintegración de proteínas y fosfolípidos en la membrana celular. Adicionalmente, el NO interviene de una manera directa en el proceso de neurodegeneración a través de la inhibición irreversible del complejo 1 (NADH deshidrogenasa) mitocondrial, mediante la nitrosilación de los grupos tioles del complejo enzimático, con lo cual se inhibe la respiración mitocondrial y, por consiguiente, la producción de ATP.26
![]()
![]()