

Interferencia por ARN (ARNi)
Cómo hemos visto existen varias alternativas para silenciar genes mediante
el uso de sistemas dependientes de la homología que funcionan en trans-.
Entre estos destacan dos modalidades: en una de ellas la interferencia ocurre
a través de procesos mediados por homología que afectan el templado,
como son las moléculas anti-sentido y formadoras de triple hélices.
En la otra modalidad, la trascripción del locus no se ve afectada,
pero la vida media de las moléculas involucradas es modificada. Tal
es el caso de la degradación post-transcripcional del ARN, mediante
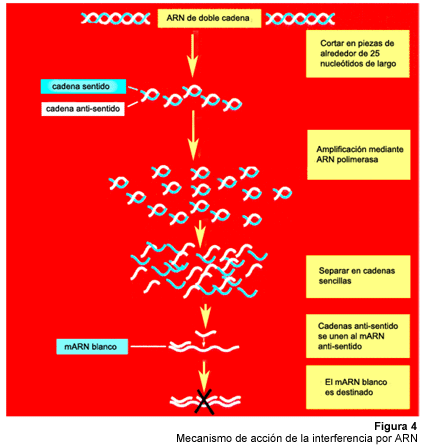
ribozimas. Recientemente, se ha demostrado que el ARN de doble cadena causa
un silenciamiento génico de manera secuencia específica, en
una gran variedad de organismos que incluyen nematodos, plantas, tripanosomas,
drosófilas y planarias4. Aún en genes que son transcritos
de manera abundante, unas cuantas moléculas de ARN por célula
pueden producir inhibición específica, lo cual no puede ser
explicado mediante un simple mecanismo de anti-sentido. La actividad sub-estequiométrica
de la interferencia por ARN (o ARNi) ha llevado a la formulación de
varios modelos: una primera opción explica que la interferencia involucra
un mecanismo catalítico dependiente del ARN inyectado; también
es factible que el material introducido sea amplificado; y, por último,
pudiese pensarse que la interferencia ocurre al nivel del gen10.
Una de las hipótesis más atractivas es que el ARN exógeno
puede dar origen a la degradación temprana del mARN endógeno,
y es posible que exista una relación entre la capacidad del ARN de
mediar la metilación del ADN, tal como se ha observado en plantas.
La importancia de este fenómeno en términos fisiológicos
no esta clara, pero se piensa que pudiera jugar un importante papel en la
modulación de la expresión génica.
![]()