
![]()
La partícula viral
El virus de la inmunodeficiencia humana (VIH), es un bien conocido ejemplo de cómo una infección viral altera la homeóstasis celular y por ende la del huésped infectado. Los virus de la inmunodeficiencia humana (VIH-1 y VIIH-2) y los virus de inmunodeficiencia simio (VISs) son los agentes etiológicos del síndrome de inmunodeficiencia adquirida (SIDA) en humanos y primates, respectivamente19,20,21,22. HIV-1, HIV-2 y SIVs pertenecen a la familia Retroviridae, virus del tipo ARN que requieren un intermediario ADN para replicación. Los retrovirus son un grupo de virus isométricos envueltos de unos 90-140 nm de diámetro y que poseen dos tipos básicos de arquitectura viral que se encuentran tanto en partículas inmaduras como en partículas maduras. Al tipo 1 de arquitectura viral pertenece los tipos B, C, D y los spumaviridae, mientras que los lentivirus y el grupo viral de la leucemia bovina/virus de la leucemia de células T humana(VLB/VLTH) poseen una arquitectura del tipo 223. VIH posee cerca de 70 espículas/virión sobre su envoltura lipídica26. Las espículas consisten de una molécula gp41 transmembranal que interactúa no-covalentemente con una mólecula gp120 formando así una estructura oligomérica de carácter trimérico27,28. La proteína de matrix (p17) delimita la membrana externa del virus de su core interno en una estructura de simetría icosadeltahédrica, y similarmente a otros lentivirus, la partícula madura de VIH contiene un core ribonucleoprotéico consistente en las proteínas p24 y p7/p625,29. Dos actividades enzimáticas se encierran en el core interno del virus, una actividad reversa transcriptasa/Rnasa H (RT) y una actividad integrasa (IN). El enzima reversa-transcriptasa permite replicar el genoma viral ARN utilizando un intermediario ADN y la actividad integrasa o endonucleasa, la cual cataliza la integración de la doble banda de ADN proviral del genoma viral en el interior del cromosoma huésped32. Los retrovirus contienen dos copias de un ARN de simple cadena/positiva de 7-10 kpb de longitud localizados en el core interno del virión. En las particulas maduras estas dos copias existen como un dímero unido no covalentemente ("kissing loop dimer")24. Un esquema de una particula viral madura se muestra en la Fig. 1.
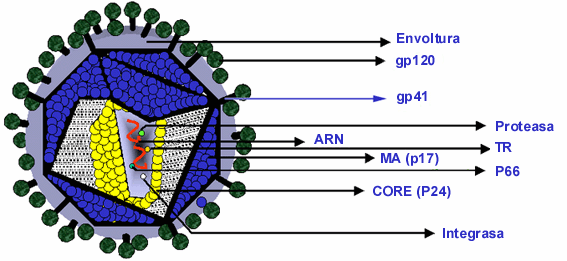
| Fig.1. Organización de una partícula madura de VIH-1. La RNP es constituida por el core viral (nucleocápside), la integrasa, la proteasa y la transcriptasa reversa(TR). Una reorganización importante del interior del virión se produce gracias a un procesamiento proteolítico de la proteína MA. Una nueva disposición geométrica de la proteína Ma esta asociada a la capacidad infecciosa. El dímero de ARN genómico se encuentra condensado en la NC. Las glicoproteínas de superficie (Gp120/gp41) se organizan sobre la envoltura viral. |
VIH al igual que otros lentivirus de primates posee un genoma complejo de 9.7 Kb en la forma de provirus y los genes estructurales retrovirales gag, pol, y env, flanqueado en las extremidades 5’ y 3’ por los LTR ("long terminal repeats"). Una aspartyl proteasa, el cual realiza el procesamiento proteolítico de los productos poliprotéicos gag y gag-pol, es también codificada por el genoma viral31. En adición a éstos genes, el genoma de VIH contiene varios genes suplementarios no encontrados en la familia Retroviridae que codifican para proteínas denominadas auxiliares (Tat, Rev, Vif, Vpr, Vpu y Nef) 5, 25. La fig. 2 muestra una representación esquemática del genoma viral, su procesamiento y sus productos proteicos.
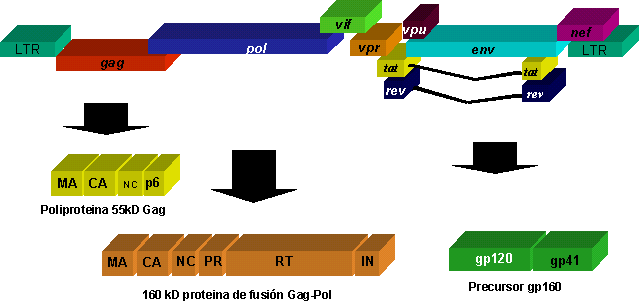
Fig. 2 Estructura genómica y procesamiento de las proteínas virales. La poliproteína meristilada de 55 kD Gag está compuesta de los dominios MA (matriz), CA (cápside), NC (nucleocápside) y p6 (con actividad facilitadora durante la liberación de la partícula y responsable de la incorporación de la proteína Vpr). Por acción de un corrimiento en el cuadro de lectura (frameshift) se produce una proteína de fusión (Gag-Pol) de unos 160 kD que provee de las actividades enzimáticas, tales reversa-transcriptas, RNasa H, proteasa. Las glicoproteínas de superficie (gp120 y gp41)son producidas a partir de un precursor poliprotéico de 160 kD y traducido a partir del gen env. El genoma de VIH codifica a su vez para varias proteínas accesorias denominadas Nef, Tat, Rev, Vif, Vpr y Vpu.
![]()
![]()