
![]()
Mecanismo de exocitosis
Aspecto
molecular
El proceso de secreción, específicamente exocitosis, involucra
varias etapas. Nosotros analizaremos con más detalle este
mecanismo a nivel de un terminal nervioso donde ya hemos descrito
los diferentes elementos involucrados como son las vesículas,
las diversas proteínas, los canales de Ca++, etc.
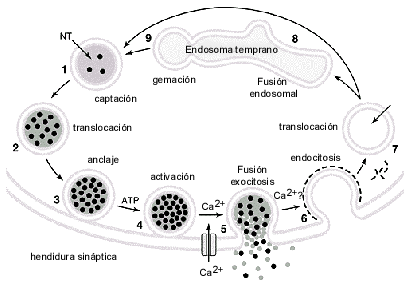
Así, una vez formada la vesícula sináptica a partir del endosoma temprano (1) para vesículas pequeñas, y/o a partir del transporte axonal rápido para las vesículas densas, se localizan en sitios cerca de las zonas activas denominados "compartimento de reserva". Allí se mantienen gracias a la interacción con la actina, porfilina u otros elementos del citoesqueleto con proteínas específicas de la membrana vesicular fundamentalmente la sinapsina y las NSF y SNAP (ver arriba). Paralelo al proceso de síntesis, transporte direccional y almacenaje de las vesículas, se realiza la síntesis de las sustancias NTs. Así los NTs clásicos como también los peptidérgicos van a ser sintetizados en el aparato de Golgi, a nivel de el cuerpo neuronal y son transportadas en vesículas a sus sitios de almacenaje. Los NTs clásicos, también van a ser sintetizados a nivel del terminal. Allí, las vesículas se van a recargar con el NT a través de los transportadores específicos. Esta captura activa del NT está dada por el gradiente electroquímico establecido por la bomba de protones vesicular. Las vesículas ya cargadas y fijadas al citoesqueleto se consideran en el "compartimento de reserva" . De este compartimento las vesículas van a ser dirigidas y ancladas específicamente por la interacción de ciertas proteínas a los sitios activos de la membrana plasmática (2-3) para este paso se considera fundamental el complejo proteico Sec6/8, munc18, sinaptobrevina, SNAP25, sintaxina forman el complejo SNAREv vesicular y se unen a los elementos SNAREt plasmáticos. Este almacén de vesículas se denomina el compartimento listo para librar (RRP: ready–releasable–pool). Una vez ancladas las vesículas van a ser activadas (4) a través de la activación de la sinaptotagmina y unión a la sintaxina. Este proceso requiere de Ca++. Además están involucradas otras proteínas como las Rabs que se unen reversiblemente al complejo SNARE y requiere GTP. Las Rabs también se han descrito involucradas en el anclaje de las vesículas.
 Todavía existe controversia en la exacta secuencia
de la cascada de activación de las diversas proteínas
involucradas en la exocitosis, como igualmente sobre cuáles
proteínas son las responsables del proceso constitutivo de
exocitosis y cuáles en el regulado.
Todavía existe controversia en la exacta secuencia
de la cascada de activación de las diversas proteínas
involucradas en la exocitosis, como igualmente sobre cuáles
proteínas son las responsables del proceso constitutivo de
exocitosis y cuáles en el regulado.
El paso de la fusión de la vesícula (5) todavía no está bien establecido. Se considera que al formarse el complejo SNARE y luego por la hidrólisis de ATP por NSF se disocia el complejo y es capaz por tensión superficial lograr la fusión de los lípidos de la membrana vesicular con la plasmática. En este paso se considera que la presencia de canales de Ca++ y su activación es fundamental. Así la sinaptotagmina activada por el Ca++ dimeriza el complejo SNARE, y lo concentra en la zona activa y así aumenta la probabilidad de fusión, se hace más reproducible y rápida ya que se ha visto que la formación del complejo SNARE por sí solo, no es suficiente para inducir la fusión. Los canales de Ca++ tipo N y P/Q se unen a la sintaxina, un componente de los SNAREt garantizando la entrada rápida y en altas concentraciones del Ca++ necesario para inducir la liberación de NTs de acción rápida . También se ha descrito un mecanismo dependiente de la Ca++-calmodulina necesario para la fase final de la exocitosis. Una vez que la vesícula se fusiona a la membrana plasmática, es recuperada por el proceso de endocitosis (6) que también requiere de varios pasos y depende de la formación de clatrina entre otros, para su activación.
![]()
![]()