

Rutas de señalización mediadas por el EGFR
El EGFR es capaz de transmitir una gran variedad de señales que pueden generar respuestas celulares tan dispares como pueden ser mitogénesis, supervivencia celular, diferenciación, prevención o inducción de apoptosis e incluso migración celular. Esta variedad de respuestas ante un mismo estímulo puede depender del tipo celular y más genéricamente de las diversas condiciones fisiológicas a las cuales estén sometidas las células. Así, en cultivos celulares estas respuestas pueden depender de la densidad celular de los cultivos, del tipo de matriz extracelular a la que estén adheridas las células o de la presencia en el medio de otros factores de crecimiento u hormonas.
Cuando el ligando extracelular se une al EGFR se produce la dimerización de éste, lo que da lugar a la activación de su tirosina quinasa y la transfosforilación de los residuos de tirosina de su extremo C-terminal (Schlessinger, 1988; Ullrich and Schlessinger, 1990; Weiss and Schlessinger, 1998). Como se comentó anteriormente, los residuos de fosfotirosina del receptor activado son reconocidos por proteínas que poseen dominios SH2 (Heldi, 1991; Koch et al., 1991; Margolis, 1992) o dominios PTB (van der Geer and Pawson, 1995). Estas proteínas pueden ser de dos tipos: proteínas adaptadoras que pueden reclutar a otras proteínas transductoras, o bien factores o enzimas directamente transductores/as que tras unirse al receptor son fosforilados por éste, pasando de un estado inactivo a otro activo. Por lo tanto, mediante estos reclutamientos y/o fosforilaciones se producen cambios conformacionales y/o cambios en la localización intracelular de estas proteínas señalizadoras, siendo así capaces éstas de transmitir sus mensajes a otros componentes de las diversas rutas intracelulares de transducción de señales (ver Fig. 1).
Dentro del amplio grupo de proteínas adaptadoras, mencionaremos como ejemplos a tres de gran importancia: la proteína Grb2 (por growth factor receptor-bound protein 2), envuelta en la activación de la proteína G monomérica Ras y la vía de las proteínas quinasas activadas por mitógenos (MAPKs, por mitogen-activated protein kinases); la proteína Grb7 (por growth factor receptor-bound protein 7); y la proteína Shc (por SH2 domain-containing protein), que actúa como un sistema auxiliar de señalización alternativo pudiendo sustituir a elementos funcionales del propio receptor.
La ruta de señalización mejor conocida en diversos organismos e iniciada por el EGFR activado es la vía de las MAPKs (Cobb, 1999). Ésta se inicia por la proteína adaptadora Grb2 que posee un domino SH2, con el que interacciona con los residuos de fosfotirosina del receptor, y dos dominios SH3 (por Src homology domain 3), con los que interacciona con factores intercambiadores de nucleótidos de guanina (GEFs, por guanine nucleotide exchange factors) tales como Sos1/2 (por Son of sevenless 1 y 2). Así, tras la formación del complejo Grb2/Sos, éste se trasloca a la membrana plasmática estimulando el intercambio de nucleótidos en Ras, transformando Ras-GDP (forma inactiva) en Ras-GTP (forma activa) (Lowenstein et al., 1992; Rozakis-Adcock et al., 1993) (ver Fig. 3). El Ras activo es capaz de interaccionar y activar a las serina/treonina quinasas Raf-1, A-Raf y B-Raf (o más genéricamente denominadas MAPKKKs, por mitogen-activated protein kinase kinase kinases) (Wood et al., 1992). Estas últimas a su vez fosforilan a las tirosina/treonina quinasas duales MEK1/2 (del acronismo MAP/ERK kinases 1 y 2, también denominadas MAPKKs, por mitogen-activated protein kinase kinases), y éstas finalmente fosforilan a las serina/treonina quinasas ERK1/2 (por extracellular-regulated kinases 1 y 2, también denominadas MAPKs). Una vez que las ERK1/2 son activadas, éstas pueden fosforilar a diferentes proteínas dianas localizadas en la membrana plasmática y en el citoplasma, dando lugar a la activación de otras vías de señalización o traslocarse al núcleo y fosforilar diversos factores de transcripción como son, entre otros, c-Myc, c-Jun, c-Fos, Elk-1 y p62TCF, produciendo así la activación o la represión transcripcional de determinados genes (Davis, 1993) (ver Fig. 4). La activación de la vía Ras/MAPK parece ser imprescindible para la proliferación celular mediada por EGF, pero no parece ser esencial para la supervivencia celular, ya que este proceso está mediado por vías independientes de Ras (Walker et al., 1998). Por otro lado, líneas celulares que expresan EGFR mutados que carecen de actividad tirosina quinasa, aunque presentan ciertas alteraciones en la transmisión de señales, sí median la activación de la vía de las MAPKs, lo que implica que mecanismos alternativos son operativos en dichas células (Campos-González and Glenney, 1992).
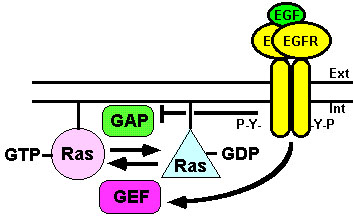
Figura 3. Regulación del ciclo
de Ras por el EGFR. El EGFR regula el ciclo de la proteína
Ras facilitando la formación de su forma activa (Ras-GTP,
círculo) mediante la activación del factor intercambiador
de nucleótidos de guanina (GEF) y secuestrando la forma
fosforilada de la proteína activadora de la GTPasa de Ras (GAP)
que la transformaría en su forma inactiva (Ras-GDP,
triángulo). Para más detalles ver texto.![]()
La proteína Grb7 contiene un dominio SH2 y pertenece a una nueva familia de proteínas adaptadoras de diversos receptores tirosina quinasa. Esta proteína se asocia a los residuos de fosfotirosina del EGFR activado y reconoce además las fosfotirosinas de los receptores homólogos ErbB2/Neu, ErbB3 y ErbB4, y las de la proteína adaptadora Shc fosforilada. Aunque las funciones de Grb7 aún no son muy bien conocidas, es indudable que juega un papel muy importante en la transducción de señales mediadas por los receptores ErbB. Así, es frecuente observar la sobreexpresión de esta proteína adaptadora junto con la de algunos de estos receptores, particularmente de ErbB2/Neu, en células de cáncer de mama (Margolis et al., 1992; Stein et al., 1994; Fiddes et al., 1998; Daly, 1998).
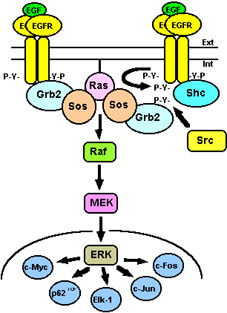
Figura 4. Activación
alternativa de la vía de la MAPK. La activación de la vía
Ras/MAPK se inicia por el complejo Grb2/Sos que es reclutado por
el EGFR transfosforilado (izquierda) o por la proteína
adaptadora Shc fosforilada previamente, bien por el propio EGFR o
por la tirosina quinasa Src (derecha). El resultado final
es la fosforilación de múltiples factores de transcripción en
el núcleo. Para más detalles ver texto.
![]()
La proteína adaptadora Shc juega un papel muy importante en la transducción de señales mediadas por el EGFR. Por un lado, esta proteína es reclutada, mediante su único dominio SH2, por los residuos de fosfotirosina del receptor activado, y por otro, también puede ser fosforilada por el receptor u otras tirosinas quinasas auxiliares, como por ejemplo Src, generando así residuos de tirosina fosforilados en la misma (Pelicci et al., 1992; Ruff-Jamison et al., 1993). Los residuos de tirosina fosforilados en Shc pueden a su vez servir como puntos de anclaje de proteínas adaptadoras, como el propio complejo Grb2/Sos, que activaría a Ras y posteriormente a la vía de las MAPKs (ver Fig. 4). Por lo tanto, células que expresan EGFR mutados que carecen de las tirosinas de su extremo C-terminal susceptibles de ser transfosforiladas, son capaces sin embargo de señalizar a través de éstos, debido a que los residuos de fosfotirosina de Shc actúan como sustitutos de los propios residuos de fosfotirosina del receptor (Li et al., 1994).
En el grupo de factores o enzimas transductores/as de la señal del EGFR mencionaremos a las proteínas inactivadoras de Ras denominadas GAPs (por GTPase-activating proteins), la fosfolipasa Cg (PLCg, por phospholipase Cg), la fosfolipasa A2 (PLA2, por phospholipase A2) y la fosfatidilinositol-3 quinasa de clase I (PI3K, por phosphatidylinositol-3 kinase).
La regulación del ciclo de intercambio de nucleótidos en Ras mediado por el EGFR, en el que participan las proteínas GEFs mencionadas anteriormente, que transforman el Ras-GDP (inactivo) en Ras-GTP (activo), se completa con la acción de las proteínas GAPs. Éstas incrementan la actividad GTPasa intrínseca de Ras, transformando de nuevo el Ras-GTP (activo) en Ras-GDP (inactivo) (ver Fig. 3). La proteína p120-GAP contiene dos dominios SH2 con los que interacciona con los residuos de fosfotirosina del EGFR activado, lo que induce la fosforilación de esta proteína moduladora (Anderson et al., 1990; Liu and Pawson, 1991). Sin embargo, esta fosforilación no altera su acción sobre la actividad GTPasa intrínseca de Ras. No obstante, la p120-GAP fosforilada se asocia firmemente al EGFR, a su homólogo ErbB2/Neu o a otras proteínas (Serth et al., 1992; Auricchio et al., 1994), por lo que esta fosforilación tiene el efecto de disminuir la disponibilidad de p120-GAP para inactivar a Ras. Por lo tanto, este mecanismo de secuestro cooperaría con la activación de Ras ejercida por las proteínas GEFs activadas a su vez por el EGFR (ver Fig. 3).
La PLCg tras interaccionar con las fosfotirosinas del EGFR activado a través de sus dos dominios SH2 es fosforilada por el receptor. Este proceso da lugar a la activación de esta enzima, la cual cataliza la hidrólisis del fosfatidilinositol 4,5-bisfosfato (PIP2, por phosphatidyl 4,5-bisphosphate) generando inositol 1,4,5-trisfosfato (IP3, por inositol 1,4,5-trisphosphate) y 1,2-diacilglicerol (DAG) (Margolis et al., 1989; Nishibe et al., 1990; Rhee and Choi, 1992; Toker, 1998). Tanto el IP3 como el DAG son potentes segundos mensajeros que están implicados en la activación de ciertas isoformas de la proteína quinasa C (PKC, por protein kinase C), bien directamente como lo hace el DAG, o bien aumentando transitoriamente la concentración de Ca2+ citosólico como lo hace el IP3, ya que éste es un efector de canales de calcio localizados en la membrana del retículo endo/sarcoplásmico (Hughes et al., 1991; Mikoshiba, 1993; Villalobo et al., 2000) (ver Fig. 5). Aunque receptores mutados que han perdido sus tirosinas transfosforilables son capaces de fosforilar a la PLCg, sin embargo, no son capaces de producir la activación de la enzima in vivo, lo cual sugiere que para transmitir la señal es necesario, además de la fosforilación de ésta, interacciones proteína-proteína (Vega et al., 1992). La activación de la PLCg por el receptor atenúa la señal mitogénica, ya que el DAG activa a la PKC y ésta retroinhibe la actividad tirosina quinasa del receptor como veremos más adelante. Por otro lado, la motilidad celular inducida por el EGF parece ser un proceso mutuamente excluyente con la proliferación celular (Chen et al., 1996).
Otra fosfolipasa que resulta fosforilada tras la activación del EGFR es la PLA2. En este caso, sin embargo, esta fosforilación es indirecta ya que tiene lugar en residuos de serina, pudiendo estar mediada por la PKC. Esta fosfolipasa también participa en la generación de la señal del calcio tras la estimulación de las células por EGF, ya que hidroliza ciertos fosfolípidos (FL) de la membrana plasmática liberando ácido araquidónico (AA), que es transformado, a través de la vía de la 5-lipoxigenasa, en leucotrieno C4 (LC4). Este último es un potente efector que abre canales de Ca2+ presentes en la membrana plasmática, contribuyendo así al aumento transitorio de la concentración de Ca2+ citosólico mediado por el receptor (Peppelenbosch et al., 1992; Schalkwijk et al., 1995; Villalobo et al., 2000) (ver Fig. 5).
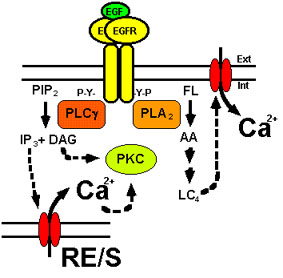
Figura 5. La señal del Ca2+
generada por el EGFR. La activación del EGFR genera un
incremento transitorio en la concentración de Ca2+ libre en el
citosol debido a la apertura de canales de Ca2+ localizados en el
retículo endo/sarcoplásmico (RE/S) y en la membrana
plasmática. Los mediadores de dicho proceso son diversos
mensajeros de naturaleza lipídica (IP3 y LC4)
sintetizados tras la activación de la PLCg y la PLA2. Las
concentraciones basales de Ca2+ se recuperan posteriormente
mediante su exclusión del citosol por diversos sistemas de
transporte (no mostrados). La acción ejercida por los
mensajeros está representada por flechas discontinuas. Para más
detalles ver texto.
![]()
Aunque la subunidad adaptadora/reguladora p85 de la PI3K de clase I contiene dominios SH2, existe controversia sobre si estos dominios son capaces o no de reconocer los residuos de fosfotirosina del EGFR activado, aunque sí parece reconocer los residuos de fosfotirosina presentes en ErbB3 (Hu et al., 1992; Soltoff et al., 1994). Así, sin excluir que la PI3K pueda interaccionar directamente con el EGFR, es posible que esta interacción se realice con una proteína adaptadora asociada a éste o bien con heterodímeros EGFR/ErbB3 (Earp et al., 1995), ya que el ErbB3 carece de actividad tirosina quinasa intrínseca, como hemos visto anteriormente, y por lo tanto sus homodímeros no son capaces de transfosforilarse (Guy et al., 1994). Una vez que la subunidad p85 de la enzima interacciona con el receptor, su subunidad catalítica p110 es activada y así es capaz de catalizar la fosforilación de la posición 3'-OH del PIP2, dando como producto de esta reacción fosfatidilinositol 3,4,5-trisfosfato (PIP3, por phosphatidyl inositol 3,4,5-trisphosphate) (Hu et al., 1992). PIP3 es un potente efector que se une a proteínas que contienen dominios PH (por pleckstrin homology domains), como es el caso de la serina/treonina quinasa PKB/Akt (PKB, por protein kinase B; y Akt, por ser homóloga de la oncoproteína v-Akt). La PKB/Akt activada fosforila a multitud de proteínas sustratos generando, entre otras, señales de supervivencia celular (Vanhaesebroeck and Waterfield, 1999). La activación de la vía PI3K-PKB/Akt también puede ocurrir por la interacción directa de Ras con la subunidad catalítica de la PI3K (Vojtek and Der, 1998), lo que previene la aparición de apoptosis (Downward, 1998). Esta vía también disminuye los niveles de expresión de la proteína supresora de la proliferación celular p53, sin que varíen los de las proteínas anti-apoptóticas Bcl-2 y Bcl-XL o los de la proteína pro-apoptótica Bax (Sabbatini and McCormick, 1999). Adicionalmente, la ruta de la PI3K-PKB/Akt induce la fosforilación de la procaspasa 9, lo que previene la formación de su forma madura activa que es fundamental en la muerte celular por apoptosis (Cardone et al., 1998) (ver Fig. 6).
Además de activar la vía ERK/MAPK, el EGFR es capaz de activar otras vías de MAPKs alternativas, como es el caso de la vía de la proteína quinasa que fosforila al factor de transcripción c-Jun (JNK, por c-Jun N-terminal kinase; también denominada SAPK, por stress-activated protein kinase). La activación de la ruta de la JNK/SAPK iniciada por el EGFR es mediada por Ras y por la PI3K (Minden et al., 1995; Coso et al., 1995; Logan et al., 1997) (ver Fig. 6). Una vez que el EGFR es activado, la proteína adaptadora Shc es fosforilada por éste, lo que permite la unión del complejo Grb2/Sos a la misma, produciéndose por tanto otra forma alternativa de activar a Ras. La interacción del EGFR con Shc parece ser imprescindible para la activación de la JNK/SAPK, mientras que no parece serlo en el caso de la activación de la vía de la ERK/MAPK, en la que el EGFR interaccionaría directamente con Grb2 (Hashimoto et al., 1999).
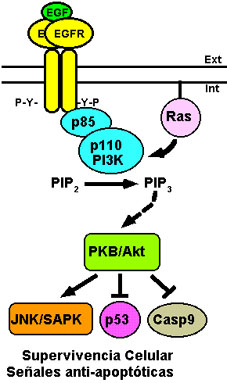
Figura 6. Vías de
señalización del EGFR por el sistema PI3K-PKB/Akt. La
activación de la PI3K por el EGFR y por Ras da lugar a la
formación de PIP3 que se une a la quinasa PKB/Akt activándola.
Esta vía estimula a la JNK/SAPK, inhibe la expresión de p53 y
previene la liberación de la forma activa de la caspasa 9 (Casp9),
lo que da lugar a la generación de señales de supervivencia
celular y prevención de apoptosis. Para más detalles ver texto.
![]()
Como hemos mencionado anteriormente, en general la activación del EGFR lleva a las células a un estado de proliferación o de supervivencia celular. En determinadas ocasiones, sin embargo, la activación del EGFR puede inducir en ciertas células tumorales una parada de la proliferación celular y la inducción de apoptosis. Estos efectos parecen estar mediados por las proteínas reguladoras de la transcripción denominadas STATs (por signal transducers and activators of transcription) que aumentan la expresión del inhibidor del ciclo celular p21WAF1/CIP1, quedando así bloqueado el mismo, y de la caspasa 1, proteasa implicada en apoptosis (Chin et al., 1996, 1997) (ver Fig. 7).
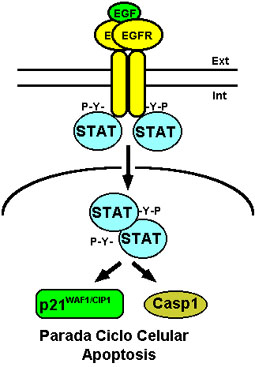
Figura 7. Inducción de apoptosis mediada por el EGFR. La fosforilación de STAT por el EGFR da lugar a la formación de dímeros fosforilados que se traslocan al núcleo e incrementan la expresión del inhibidor del ciclo celular p21WAF1/CIP1 y de la caspasa 1 (Casp1). Para más detalles ver texto.
![]()